
User:WillHHudson/Sandbox
| Substantia nigra | |
|---|---|
 Section through superior colliculus showing path of oculomotor nerve. | |
 Coronal slices of human brain showing the basal ganglia, globus pallidus: external segment (GPe), subthalamic nucleus (STN), globus pallidus: internal segment (GPi), and substantia nigra (SN, red). The right section is the deeper one, closer to the back of the head. | |
| Details | |
| Part of | Midbrain, Basal ganglia |
| Anatomical terms of neuroanatomy | |
The substantia nigra is a brain structure located in the mesencephalon (midbrain) that plays an important role in reward, addiction, and movement. Substantia nigra is Latin for "black substance", as parts of the substantia nigra appear black due to high levels of melanin in dopaminergic neurons. The substantia nigra is part of the basal ganglia; the other parts of the basal ganglia include the striatum (caudate nucleus and putamen), nucleus accumbens, and the subthalamic nucleus. Parkinson's disease is caused by the death of dopaminergic neurons in the substantia nigra pars compacta.
Anatomy[edit]
The substantia nigra, along with four other nuclei, is part of the basal ganglia. The substantia nigra lies in the midbrain, dorsal to the cerebral peduncles. Humans have two substantiae nigrae, one on each side of the midline. The substantia nigra is the largest nucleus in the midbrain. The substantia nigra is divided into two parts: the pars reticulata and pars compacta, which lies medial to the pars reticulata. Sometimes a third region, the pars lateralis, is mentioned; however, this is usually classified as part of the pars reticulata. The pars reticulata and the internal globus pallidus are separated by the internal capsule. Segment M1 of the middle cerebral artery supplies blood to all of the basal ganglia, including the substantia nigra. The neurons of the pars compacta are restricted to input from and output to the basal ganglia, while the neurons of the pars reticulata send and receive information from various parts of the brain, not necessarily the basal ganglia.
Neurons in the pars compacta are generally dopaminergic. They are sometimes referred to as the A9 cell group, the A8 and A10 cell groups being the other major dopaminergic groups in the brain, in the retrorubral area and ventral tegmental area, respectively. The dopaminergic neurons in the pars compacta can be distinguished by their blackened appearance due to a buildup of melanin. The neurons in the pars reticulata are generally GABAergic. The neurons in the substantia nigra can be classified into two broad categories: interneurons and projection neurons.[1] Interneurons connect various neurons within the substantia nigra, and projection neurons extend to other parts of the brain (usually the basal ganglia). Many projection neurons in the pars compacta send dendrites into the pars reticulata; this is one mechanism by which GABAergic input can modulate dopamine activity. [2]
Afferent Connections[edit]
The chief afferent projection into the substantia nigra is the striatonigral projection. Most striatonigral neurons terminate in the pars reticulata. Additional nigral inputs project from the globus pallidus[3], amygdala,[4] reticular formation[5], and the subthalamic nucleus[6]. GABA is the primary neurotransmitter for these connections. The amygdalo-nigrostriatal pathway has recently been implicated in surprised-induced learning.[7] All connections from non-basal ganglia nuclei project into the pars reticulata.
Efferent Projections[edit]
The most well-known and important efferent projection of the substantia nigra is the efferent striatonigral projection (sometimes called the nigrostriatal projection), which originates chiefly in the pars compacta, but also includes a smaller number of other dopaminergic cells scattered throughout the pars reticulata. The efferent striatonigral projection uses dopamine as its primary transmitter. Within the striatum, acetylcholine is the primary neurotransmitter, and the striatal dopamine-acetylcholine relationship is very important one: striatal output neurons receive inputs from the entire cerebral cortex and select relevant cortical information, and dopaminergic input from the pars compacta modulates the striatal output. In addition to their chemical synapses, there are also electrical synapses between nigrostriatal dopaminergic neurons and striatal output neurons.[8] Striatal aceytlcholine is elevated in Parkinson's disease, and the interactions between striatal acetylcholine and dopamine are critical in motor response selection, especially stimulus-reaction associations or acquired through positive reinforcement. [9]Nicotine alters this balance, which facilitates addiction.[10]
In addition, there are significant projections to the thalamus (ventral lateral and ventral anterior nuclei), superior colliculus, and other caudal nuclei from the pars reticulata (the nigrothalamic pathway).[11] These neurons use GABA as their neurotransmitter. In addition, these neurons form up to five collaterals which branch within both the pars compacta and pars reticulata, likely modulating dopaminergic activity in the pars compacta[12] .
Function[edit]
The substantia nigra is an important player in brain function, particularly in eye movement, motor planning, reward seeking, learning, and addiction. Many of the substantia nigra's effects are mediated through the striatum. The nigral dopaminergic input to the striatum via the nigrostriatal pathway is intimately linked with the striatum's function.[13] The co-dependence between the striatum and substantia nigra can be seen in this way: when the substantia nigra is electrically stimulated, no movement occurs; however, the symptoms of nigral degeneration due to Parkinson's is a poignant example of the substantia nigra's influence on movement. In addition to striatum-mediated functions, the substantia nigra also serves as a major sources of GABAergic inhibition to various brain targets.
Pars Reticulata[edit]
The pars reticulata of the substantia nigra is an important processing center in the basal ganglia. The GABAergic neurons in the pars reticulata convey the final processed signals of the basal ganglia to the thalamus and superior colliculus. In addition, the pars reticulata also inhibits dopaminergic activity in the pars compacta via axon collaterals, although the functional organization of these connections remains unclear.
The GABAergic neurons of the pars reticulata spontaneously fire action potentials. In rats, the frequency of action potentials is roughly 25Hz.[14] The purpose of these spontaneous action potentials is to inhibit targets of the basal ganglia, and decreases in inhibition are associated with movement.[15] The subthalamic nucleus gives excitatory input that modulates the rate of firing of these spontaneous action potentials. However, lesion of the subthalamic nucleus leads to only a 20% decrease in pars reticulata firing rate, suggesting that the generation of action potentials in the pars reticulata is largely autonomous.[16] An example of this inhibitory output is the important role of the pars reticulata in saccadic eye movement. A group of GABAergic neurons from the pars reticulata projects to the superior colliculus. This connection exhibits a high level of sustained, inhibitory activity.[17] Projections from the caudate nucleus to the superior colliculus also modulate saccadic eye movement. Altered patterns of pars reticulata firing such as single-spike or burst firing are found in Parkinson's disease[18] and epilepsy.[19]
Pars Compacta[edit]
The most prominent function of the pars compacta is motor control.[20] However, the substantia nigra's role in motor control is indirect; electrical stimulation of the substantia nigra does not result in movement. This is due to the mediation of the striatum in the nigral influence of movement. However, lack of pars compacta neurons clearly has a large influence on movement, as evidenced by the symptoms of Parkinson's. The motor role of the pars compacta may involve fine motor control; this has been confirmed in animal models with lesions in the pars compacta.[21]
The pars compacta is heavily involved in learned responses to stimuli. In primates, dopaminergic neuron activity increases in the nigrostriatal pathway when a new stimulus is presented.[22] Dopaminergic activity decreases with repeated stimulus presentation.[22] However, behaviorally significant stimulus presentation (such as classical conditioning where a reward is presented) continues to activate the dopaminergic neurons. This phenomenon helps explain the role of the dopamine system in the addictiveness of drugs. In addition, the pars compacta is important in "spatial learning," the observations about one's environment and location in space. Lesions in the pars compacta lead to learning deficits in repeating identical movements[23], and some studies point to a possible spatial memory system independent from the hippocampus, where spatial memory is traditionally thought to be located.[24]
Temporal processing is also an important function of the pars compacta. The pars compacta is activated during time reproduction and lesions in the pars compacta leads to temporal deficits.[25] As of late, the pars compacta has been suspected of regulating the sleep-wake cycle.[26] This is consistent with symptoms such as insomnia and REM sleep disturbances that are reported by patients with Parkinson's disease. Even so, partial dopamine deficits that do not affect motor control can lead to disturbances in the sleep-wake cycle, especially REM-like patterns of neural activity while awake, especially in the hippocampus.[27]
Pathophysiology[edit]
The substantia nigra is critical in the development of many diseases, including Parkinson's disease.
Parkinson's Disease[edit]
Parkinson's disease's is a neurodegenerative disease caused by the death of dopaminergic neurons in the pars compacta of the substantia nigra. The major symptoms of Parkinson's disease include tremor, akinesia, bradykinesia, and stiffness.[28] Other symptoms include disturbances to posture, fatigue, sleep abnormalities, and depression.[29]
The cause of death of dopaminergic neurons in the pars compacta is unknown. However, some contributions to the unique susceptibility of dopaminergic neurons in the pars compacta have been identified. For one, dopaminergic neurons show abnormalities in mitochondrial complex 1, causing aggregation of alpha-synuclein. This could result in abnormal protein handling and neuron death.[30] Secondly, dopaminergic neurons in the pars compacta contain less calbindin than other dopaminergic neurons.[31] Calbindin is a protein involved in calcium ion transport within cells, and excess calcium in cells is toxic. The calbindin theory would explain the high cytotoxicity of Parkinson's in the substantia nigra compared to the ventral tegmental area. Regardless of the cause of neuronal death, the plasticity of the pars compacta is very robust; Parkinsonian symptoms do not appear until up to 50-80% of pars compacta dopaminergic neurons have died.[32] Most of this plasticity occurs at the neurochemical level; dopamine transport systems are slowed, allowing dopamine to linger for longer periods of time in the chemical synapses in the striatum.[32]
Schizophrenia[edit]
Increased levels of dopamine have long been implicated in the development of schizophrenia.[33] However, much debate continues to this day surrounding this theory, commonly known as the dopamine hypothesis of schizophrenia. Despite the controversy, dopamine antagonists remain a standard and successful treatment for schizophrenia. These antagonists include first generation (typical) antipsychotics such as butyrophenones, phenothiazines, and thioxanthenes. These drugs have largely been replaced by second generation (atypical) antipsychotics such as clozapine and paliperidone. It should be noted that these drugs generally do not act on dopamine-producing neurons themselves, but on the dopaminergic receptors on which the dopaminergic neurons synapse.
Other, non-pharmacological evidence in support of the dopamine hypothesis relating to the substantia nigra include structural changes in the pars compacta, such as reduction in synaptic terminal size.[34] Other changes in the substantia nigra include increased expression of NMDA receptors in the substantia nigra, and reduced dysbindin expression. Increased NMDA receptors may point to the involvement of glutamate-dopamine interactions in schizophrenia. Dysbindin, which has been (controversially) linked to schizophrenia, may regulate dopamine release, and low expression of dysbindin in the substantia nigra may be important in schizophrenia etiology.[35] Due to the changes to the substantia nigra in the schizophrenic brain, it may eventually be possible to use specific imaging techniques (such as melanin-specific imaging) to detect physiological signs of schizophrenia in the substantia nigra.[36]
Chemical Modification of the Substantia Nigra[edit]
Chemical manipulation and modification of the substantia nigra is important in the fields of neuropharmacology and toxicology. Various compounds such as levodopa and MPTP are used in the treatment and study of Parkinson's disease, and many other drugs have effects on the substantia nigra.
Levodopa[edit]
The substantia nigra is the target of chemical therapeutics for the treatment of Parkinson's disease. Levodopa (L-DOPA), the dopamine precursor, is the most commonly prescribed medication for Parkinson's disease. Despite controversy concerning the neurotoxicity [37] of dopamine, it remains the most common treatment for Parkinson's disease. The drug is especially effective in treating patients in the early stages of Parkinson's, although the drug does lose its efficacy over time.[38] Levodopa can cross the blood-brain barrier, and increases dopamine levels in the substantia nigra, thus alleviating the symptoms of Parkinson's disease. The drawback of levodopa treatment is that it treats the symptoms of Parkinson's (low dopamine levels), rather than the cause (the death of dopaminergic neurons in the substantia nigra).
MPTP[edit]
MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine), is a neurotoxin specific to dopaminergic cells in the brain, specifically in the substantia nigra. MPTP was brought to the spotlight in 1982 when heroin users in California displayed Parkinson's-like symptoms after using heroin contaminated with MPTP. The patients bodies were rigid nearly completely immobile, and they responded to levodopa treatment. No remission of the Parkinson's-like symptoms was reported, suggesting irreversible death of the dopaminergic neurons.[39] The proposed mechanism of MPTP involves disruption of mitochondial function, including disruption of metabolism and creation of free radicals.[40]
Soon after, MPTP was tested in animal models for its efficacy in inducing Parkinson's disease (with success). MPTP induced akinesia, rigidity, and tremor in primates, and its neurotoxicity was found to be very specific to the substantia nigra pars compacta.[41] In other animals, such as rodents, the induction of Parkinson's by MPTP is incomplete or requires much higher and frequent doses than in primates. Today, MPTP remains the most favored model for studying Parkinson's. [40]
Cocaine[edit]
Cocaine's mechanism of action in the human brain includes the inhibition of dopamine reuptake.[42] This blockage of dopamine reuptake explains cocaine's addictive properties, as dopamine is the critical neurotransmitter for reward. However, cocaine is more active in the dopaminergic neurons of the ventral tegmental area than the substantia nigra. Cocaine administration increases metabolism in the substantia nigra, which can explain the altered motor function seen in cocaine-using subjects.[43] The inhibition of dopamine reuptake by cocaine also inhibits the firing of spontaneous action potentials by the pars compacta.[44] The mechanism by which cocaine inhibits cocaine reuptake involves its binding to the dopamine transporter protein, or DAT. However, recent studies show that cocaine can also cause a decrease in DAT mRNA levels.[45] This is probably caused by cocaine's blockade of DAT rather than direct interference with transcriptional or translational pathways.[45]
Inactivation of the substantia nigra could prove to be a possible treatment for cocaine addiction. In a study of cocaine-dependent rats, inactivation of the substantia nigra via implanted cannulae greatly reduced cocaine addiction relapse.[46]
Amphetamines[edit]
Like cocaine, amphetamine increases the concentrations of dopamine in the synaptic cleft, thereby heightening the response of the post-synaptic neuron.[47] Also like cocaine, the altered dopamine function contributes to the addictiveness of amphetamines.
The specific mechanisms by which amphetamines affect dopamine concentrations is not well understood. Currently, two major hypotheses have been proposed. The first theory emphasizes amphetamine’s actions on the vesicular level, increasing concentrations of dopamine in the cytosol of the pre-synaptic neuron.[48] The other hypothesized mechanism, whichis similar to that of cocaine's, focuses on the role of the dopamine transporter, and proposes that amphetamine may interact with DAT to induce reverse transport of dopamine from the presynaptic neuron into the synaptic cleft.[49][50]
Additional images[edit]
-
 Schematic representation of the chief ganglionic categories (I to V).
Schematic representation of the chief ganglionic categories (I to V). -
 Deep dissection of brain-stem. Lateral view.
Deep dissection of brain-stem. Lateral view. -
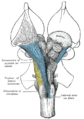 Deep dissection of brain-stem. Ventral view.
Deep dissection of brain-stem. Ventral view. -
 Coronal section through mid-brain.
Coronal section through mid-brain. -
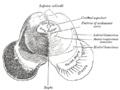 Transverse section of mid-brain at level of inferior colliculi.
Transverse section of mid-brain at level of inferior colliculi. -
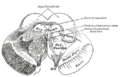 Transverse section of mid-brain at level of superior colliculi.
Transverse section of mid-brain at level of superior colliculi. -
 Coronal section of brain immediately in front of pons.
Coronal section of brain immediately in front of pons. -
 Human brain frontal (coronal) section
Human brain frontal (coronal) section -
 Dopamine and serotonin
Dopamine and serotonin







_section_description_2.JPG)

References[edit]
- ^ Danner, H; Pfister, C (1982). "7 types of neurons in the substantia nigra of the rat. Golgi rapid-impregnation study". J Hirnforsch. 23 (5): 553–6. PMID 6186738.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Juraska, J M; Wilson, C J; Groves, P M (1970). "The substantia nigra of the rat: a Golgi study". J Comp Neurol. 172 (4): 585–600. doi:10.1002/cne.901720403. PMID 65369.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Hattori, T; Fibiger, H C; McGeer, P L (1975). "Demonstration of a pallido-nigral projection innervating dopaminergic neurons". J Comp Neurol. 162 (4): 487–504. doi:10.1002/cne.901620406. PMID 50334.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Han, J S; McMahan, R W; Holland, P; Gallagher, M (1997). "The role of an amygdalo-nigrostriatal pathway in associative learning". J Neurosci. 17 (10): 3913–9. doi:10.1523/JNEUROSCI.17-10-03913.1997. PMC 6573694. PMID 9133409.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ "Demonstration of axon collateral projections from the substantia nigra pars reticulata to the superior colliculus and the parvicellular reticular formation in the rat". Brain Res. 674 (1): 122–126. 1995. doi:10.1016/0006-8993(94)01459-U. PMID 7539705.
{{cite journal}}: Cite uses deprecated parameter|authors=(help) - ^ Nauta, H J; Cole, M (1978). "Efferent projections of the subthalamic nucleus: an autoradiographic study in monkey and cat". J Comp Neurol. 180 (1): 1–16. doi:10.1002/cne.901800102. PMID 418083.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Lee, H J; Youn, J M; Gallagher, M; Holland, P C (2008). "Temporally limited role of substantia nigra-central amygdala connections in surprise-induced enhancement of learning". Eur J Neurosci. 27 (11): 3043–3049. doi:10.1111/j.1460-9568.2008.06272.x. PMC 2853798. PMID 18588542.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Vandecasteele, M; et al. (2004). Electrical Synapses Between Output Neurons of the Striatum and Between Neurons of the Substantia Nigra Pars Compacta. Basal Ganglia VIII. Springer Science+Business Media. pp. 493–502. ISBN 0387280650.
{{cite book}}: Explicit use of et al. in:|author=(help) - ^ Aosaki, T; Tsubokawa, H; Ishida, A; Watanabe, K; Graybiel, A M; Kimura, M (1994). "Responses of tonically active neurons in the primate's striatum undergo systematic changes during behavioral sensorimotor conditioning". J Neurosci. 14 (6): 3969–84. doi:10.1523/JNEUROSCI.14-06-03969.1994. PMC 6576948. PMID 8207500.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Rice, Margaret E; Cragg, Stephanie J (2004). "Nicotine amplifies reward-related dopamine signals in striatum". Nat Neurosci. 7 (6): 583–4. doi:10.1038/nn1244. PMID 15146188.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Carpenter, M B; Nakano, K; Kim, R (1976). "Nigrothalamic projections in the monkey demonstrated by autoradiographic technics". J Comp Neurol. 165 (4): 401–15. doi:10.1002/cne.901650402. PMID 57125.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Deniau, J M; Kitai, S T; Donoghue, J P; Grofova, I (1992). "Neuronal interactions in the substantia nigra pars reticulata through axon collaterals of the projection neurons. An electrophysiological and morphological study". Exp Brain Res. 47 (1): 105–13. doi:10.1007/BF00235891. PMID 6288427.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Nicola, S M; Surmeier, J; Malenka, R C (2000). "Dopaminergic modulation of neuronal excitability in the striatum and nucleus accumbens". Annu Rev Neurosci. 23: 185–215. doi:10.1146/annurev.neuro.23.1.185. PMID 10845063.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Gernert, Manuela; Fedrowitz, Maren; Wlaz, Piotr; Loscher, Wolfgang (2004). "Subregional changes in discharge rate, pattern, and drug sensitivity of putative GABAergic nigral neurons in the kindling model of epilepsy". Eur J Neurosci. 20 (9): 2377–2386. doi:10.1111/j.1460-9568.2004.03699.x. PMID 15525279.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Sato, Makoto; Hikosaka, Okihide (2002). "Role of primate substantia nigra pars reticulata in reward-oriented saccadic eye movement". J Neurosci. 22 (6): 2363–2373. doi:10.1523/JNEUROSCI.22-06-02363.2002. PMC 6758246. PMID 11896175.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Zahr, Natalie May; Martin, Lynn Pauline; Waszczak, Barbara Lee (2004). "Subthalamic nucleus lesions alter basal and dopamine agonist stimulated electrophysiological output from the rat basal ganglia". Synapse. 53 (2): 119–28. doi:10.1002/syn.20064. PMID 15352137.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Hikosaka, O; Wurtz, R H (1983). "Visual and oculomotor functions of monkey substantia nigra pars reticulata. III. Memory-contingent visual and saccade responses". J Neurophysiol. 49 (5): 1268–84. doi:10.1152/jn.1983.49.5.1268. PMID 6864250.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Tseng, K Y; Riquelme, L A; Belforte, J E; Pazo, J H; Murer, M G (2000). "Substantia nigra pars reticulata units in 6-hydroxydopamine-lesioned rats: responses to striatal D2 dopamine receptor stimulation and subthalamic lesions". Eur J Neurosci. 12 (1): 247–256. doi:10.1046/j.1460-9568.2000.00910.x. PMID 10651879.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Deransart, Colin; Hellwig, Bernhard; Heupel-Reuter, Miriam; Leger, Jean-Francois; Heck, Detlef; Lucking, Carl Hermann (2003). "Single-unit analysis of substantia nigra pars reticulata neurons in freely behaving rats with genetic absence epilepsy". Epilepsia. 44 (12): 1513–20. doi:10.1111/j.0013-9580.2003.26603.x. PMID 14636321.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Hodge, G K; Butcher, L L (1980). "Pars compacta of the substantia nigra modulates motor activity but is not involved importantly in regulating food and water intake". Naunyn Schmiedebergs Arch Pharmacol. 313 (1): 51–67. doi:10.1007/BF00505805. PMID 7207636.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Pioli, E Y; Meissner, W; Sohr, R; Gross, C E; Bezard, E; Bioulac, B H (2008). "Differential behavioral effects of partial bilateral lesions of ventral tegmental area or substantia nigra pars compacta in rats". Neuroscience. 153 (4): 1213–24. doi:10.1016/j.neuroscience.2008.01.084. PMID 18455318.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ a b Ljungberg, T; Apicella, P; Schultz, W (1992). "Responses of monkey dopamine neurons during learning of behavioral reactions". J Neurophysio. 67 (1): 145–163. doi:10.1152/jn.1992.67.1.145. PMID 1552316.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Da Cunha, Claudio; Silva, Marcio H C; Wietzikoski, Samantha; Wietzikoski, Evellyn C; Ferro, Marcelo M; Kouzmine, Ivana; Canteras, Newton S (2006). "Place learning strategy of substantia nigra pars compacta-lesioned rats". Behav Neurosci. 120 (6): 1279–1284. doi:10.1037/0735-7044.120.6.1279. PMID 17201473.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Da Cunha, Claudio; Wietzikoski, Samantha; Wietzikoski, Evellyn C; Miyoshi, Edmar; Ferro, Marcelo M; Anselmo-Franci, Janete A; Canteras, Newton S (2003). "Evidence for the substantia nigra pars compacta as an essential component of a memory system independent of the hippocampal memory system". Neurobiol Learn Mem. 79 (3): 236–242. doi:10.1016/S1074-7427(03)00008-X. PMID 12676522.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Matell, M S; Meck, W H (2000). "Neuropsychological mechanisms of interval timing behavior". BioEssays. 22 (1): 94–103. doi:10.1002/(SICI)1521-1878(200001)22:1<94::AID-BIES14>3.0.CO;2-E. PMID 10649295.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Lima, Marcelo M S; Andersen, Monica L; Reksidler, Angela B; Vital, Maria A B F; Tufik, Sergio (2007). "The role of the substantia nigra pars compacta in regulating sleep patterns in rats". PLoS ONE. 2 (6): e513. doi:10.1371/journal.pone.0000513. PMC 1876809. PMID 17551593.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Dzirasa, Kafui; Ribeiro, Sidarta; Costa, Rui; Santos, Lucas M; Lin, Shih-Chieh; Grosmark, Andres; Sotnikova, Tatyana D; Gainetdinov, Raul R; Caron, Marc G; Nicolelis, Miguel A L (2006). "Dopaminergic control of sleep-wake states". J Neurosci. 26 (41): 10577–10589. doi:10.1523/JNEUROSCI.1767-06.2006. PMC 6674686. PMID 17035544.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Jankovic J (April 2008). "Parkinson's disease: clinical features and diagnosis". J. Neurol. Neurosurg. Psychiatr. 79 (4): 368–76. doi:10.1136/jnnp.2007.131045. PMID 18344392.
{{cite journal}}: CS1 maint: date and year (link) - ^ Adler, CH (2005). "Nonmotor complications in Parkinson's disease". Movement Disorders. 20: S23-9. doi:10.1002/mds.20460. PMID 15822106.
- ^ Dawson, T; Dawson, V (2003). "Molecular Pathways of Neurodegeneration in Parkinson's Disease". Science. 302 (5646): 819–822. doi:10.1126/science.1087753. PMID 14593166.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Liang CL, Sinton CM, Sonsalla PK, German DC (1996). "Midbrain dopaminergic neurons in the mouse that contain calbindin-D28k exhibit reduced vulnerability to MPTP-induced neurodegeneration". Neurodegeneration. 5 (4): 313–318. doi:10.1006/neur.1996.0042. PMID 9117542.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ a b Interview. Yoland Smith, PhD
- ^ van Rossum, J (1967). Brill H, Cole J, Deniker P, Hippius H, Bradley P B (ed.). Neuropsychopharmacology, Proceedings Fifth Collegium Internationale Neuropsychopharmacologicum: 321-9.
{{cite journal}}: Missing or empty|title=(help)CS1 maint: multiple names: editors list (link) - ^ N. S. Kolomeets and N. A. Uranova (1997). "Synaptic contacts in schizophrenia: Studies using immunocytochemical identification of dopaminergic neurons". Neuroscience and Behavioral Physiology: 217-221.
- ^ Kumamoto, N.; Matsuzaki, S.; Inoue, K.; Hattori, T.; Shimizu, S.; Hashimoto, R.; Yamatodani, A.; Katayama, T.; Tohyama, M. (2006). "Hyperactivation of midbrain dopaminergic system in schizophrenia could be attributed to the down-regulation of dysbindin". Biochem Biophys Res Commun. 345 (2): 904–909. doi:10.1016/j.bbrc.2006.04.163. PMID 16701550.
- ^ Shibata, Eri; Sasaki, Makoto; Tohyama, Koujiro; Otsuka, Kotaro; Endoh, Jin; Terayama, Yasuo; Sakai, Akio (2008). "Use of neuromelanin-sensitive MRI to distinguish schizophrenic and depressive patients and healthy individuals based on signal alterations in the substantia nigra and locus ceruleus". Biol Psychiatry. 64 (5): 401–406. doi:10.1016/j.biopsych.2008.03.021. PMID 18452894.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Cheng N, Maeda T, Kume T; et al. (December 1996). "http://www.ncbi.nlm.nih.gov/pubmed/9017256". Brain Research. 743 (1–2): 278–83. doi:10.1016/S0006-8993(96)01056-6. PMID 9017256.
{{cite journal}}: Explicit use of et al. in:|author=(help); External link in|title= - ^ Rascol O, Payoux P, Ory F, Ferreira JJ, Brefel-Courbon C, Montastruc JL. (2003). "Limitations of current Parkinson's disease therapy". Ann Neurol. 53 (S3): S3–S15. doi:10.1002/ana.10513. PMID 12666094.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ JW Langston, P Ballard, JW Tetrud, and I Irwin (1983). "Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis". Science. 219 (4587): 979–980. doi:10.1126/science.6823561. PMID 6823561.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ a b Schmidt, N; Ferger, B (2001). "Neurochemical findings in the MPTP model of Parkinson's disease". J Neural Transm. 108 (11): 1263–82. doi:10.1007/s007020100004. PMID 11768626.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Langston, J W; Forno, L S; Rebert, C S; Irwin, I (1984). "Selective nigral toxicity after systemic administration of 1-methyl-4-phenyl-1,2,5,6-tetrahydropyrine (MPTP) in the squirrel monkey". Brain Res. 292 (2): 390–394. doi:10.1016/0006-8993(84)90777-7. PMID 6607092.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Heikkila, R E; Cabbat, F S; Duvoisin, R C (1979). "Motor activity and rotational behavior after analogs of cocaine: correlation with dopamine uptake blockade". Commun Psychopharmacol. 3 (5): 285–90. PMID 575770.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Joan M. Lakoski, Matthew P. Galloway, Francis J. White (1991). Cocaine. Telford Press. ISBN 0849388139.
{{cite book}}: CS1 maint: multiple names: authors list (link) - ^ M. G. Lacey, N. B. Mercuri, and R. A. North (1990). "Actions of cocaine on rat dopaminergic neurones in vitro". Br J Pharmacol. 99 (4): 731–735. doi:10.1111/j.1476-5381.1990.tb12998.x. PMC 1917549. PMID 2361170.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ a b Yue Xia, Dennis J. Goebel, Gregory Kapatos Michael J. Banno (2006). "Quantitation of Rat Dopamine Transporter mRNA: Effects of Cocaine Treatment and Withdrawal". J Neurochem. 59 (3): 1179–1182.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ See, R E; Elliott, J C; Feltenstein, M W (2007). "The role of dorsal vs ventral striatal pathways in cocaine-seeking behavior after prolonged abstinence in rats". Psychopharmacology (Berl). 194 (3): 321–31. doi:10.1007/s00213-007-0850-8. PMID 17589830.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Kuczenski, R and Segal, D. (1997). "Effects of methylphenidate on extracellular dopamine, serotonin, and norepinephrine; Comparison with amphetamine". J Neurochem. 68 (5): 2032–2037. doi:10.1046/j.1471-4159.1997.68052032.x. PMID 9109529.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Rothman, R; Baumann, M. (2006). "Balance between dopamine and serotonin release modulates behavioral effects of amphetamine-type drugs". Ann N Y Acad Sci. 1074 (1074): 245-60. doi:10.1196/annals.1369.064. PMID 17105921.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ Johnson, L’Aurelle A.; et al. (2005). "Regulation of amphetamine-stimulated dopamine efflux by Protein Kinase C-beta". J. Biol. Chem. 280 (12): 10914–10919. doi:10.1074/jbc.M413887200. PMID 15647254.
{{cite journal}}: Explicit use of et al. in:|author=(help) - ^ Kahlig, Kristopher et al. “Amphetamine induces dopamine efflux through a dopamine transporter channel. |url = http://www.pnas.org/content/102/9/3495.abstract” PNAS 102 (2005): 3495-3500}}